
- Home
- About
- What is a Tipping Point?
- Tipping Points Conference
- Resources
- Contact
- Summary Report
- Introduction
- Summary Report
-
-
- Summary
Harmful tipping points in the natural world pose some of the gravest threats faced by humanity. Their triggering will severely damage our planet’s life-support systems and threaten the stability of our societies.
In the Summary Report:
• Narrative summary
• Global tipping points infographic
• Key messages
• Key Recommendations
Executive summary
• Section 1
• Section 2
• Section 3
• Section 4
-
- Downloads
Summary
Report
-
Full
Report
-
Key
Messages
-
Key
Recommendations
-
Global tipping points
Infographic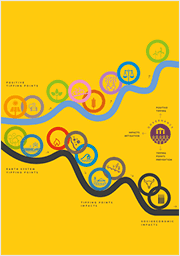
-
-
- Introduction
-
-
- Why we need to talk about tipping points
This report is for all those concerned with tackling escalating Earth system change and mobilising transformative social change to alter that trajectory, achieve sustainability and promote social justice.
In this section:
• Foreword
• Introduction
• Key Concepts
• Approach
• References
-
-
- Section 1
-
-
Considers Earth system tipping points. These are reviewed and assessed across the three major domains of the cryosphere, biosphere and circulation of the oceans and atmosphere. We then consider the interactions and potential cascades of Earth system tipping points, followed by an assessment of early warning signals for Earth system tipping points.
-
-
- 1.1 Introduction
- 1.2 Tipping points in the cryosphere
- 1.3 Tipping points in the biosphere
- 1.4 Tipping points in ocean and atmosphere circulations
- 1.5 Climate tipping point interactions and cascades
- 1.5.1 Introduction and definition
- 1.5.2 Interactions between climate tipping systems and further nonlinear climate components
- 1.5.2.1 Interactions across scales in space and time
- 1.5.2.2 Interactions between ice sheets and AMOC
- 1.5.2.3 Arctic sea ice interactions
- 1.5.2.4 Effects of AMOC changes on the Amazon rainforest
- 1.5.2.5 Interactions between ENSO and tipping systems
- 1.5.2.6 Effects of permafrost thaw on the global hydrological cycle
- 1.5.3 Archetypal examples of interactions between tipping systems from a palaeoclimatic perspective
- 1.5.4 Interactions between tipping systems and planetary-scale cascades
- 1.5.5 Final remarks
- 1.6 Early warning signals of Earth system tipping points
- 1.7 Earth system tipping points – synthesis
- References
-
-
-
- 1. Earth System Tipping Points
- 1.1 Introduction
- 1.2 Tipping points in the cryosphere
- 1.3 Tipping points in the biosphere
- 1.4 Tipping points in ocean and atmosphere circulations
- 1.5 Climate tipping point interactions and cascades
- 1.5.1 Introduction and definition
- 1.5.2 Interactions between climate tipping systems and further nonlinear climate components
- 1.5.2.1 Interactions across scales in space and time
- 1.5.2.2 Interactions between ice sheets and AMOC
- 1.5.2.3 Arctic sea ice interactions
- 1.5.2.4 Effects of AMOC changes on the Amazon rainforest
- 1.5.2.5 Interactions between ENSO and tipping systems
- 1.5.2.6 Effects of permafrost thaw on the global hydrological cycle
- 1.5.3 Archetypal examples of interactions between tipping systems from a palaeoclimatic perspective
- 1.6 Early warning signals of Earth system tipping points
- 1.7 Earth System Tipping Points – Synthesis
- References
- Section 2
-
-
Considers tipping point impacts. First we look at the human impacts of Earth system tipping points, then the potential couplings to negative tipping points in human systems. Next we assess the potential for cascading and compounding systemic risk, before considering the potential for early warning of impact tipping points.
-
-
- 2.1 Introduction
- 2.2 Assessing Impacts of Earth System Tipping Points on Human Societies
- 2.2.1 Introduction
- 2.2.2 Impacts of cryosphere tipping points
- 2.2.3 Impacts of biosphere tipping points
- 2.2.4 Impacts of ocean-atmosphere circulation tipping points
- 2.2.5 Potential for Earth system tipping points to magnify or accelerate impacts of global warming
- 2.2.6 Sector-based impacts assessment of climate system tipping points
- 2.3 Negative social tipping points
- 2.4 Cascades of tipping in impacts
- 2.5 Early warning of tipping points in impacts
- References
-
-
-
- 2. Tipping point impacts
- 2.1 Introduction
- 2.2 Assessing Impacts of Earth System Tipping Points on Human Societies
- 2.2.1 Introduction
- 2.2.2 Impacts of cryosphere tipping points
- 2.2.3 Impacts of biosphere tipping points
- 2.2.4 Impacts of ocean-atmosphere circulation tipping points
- 2.2.5 Potential for Earth system tipping points to magnify or accelerate impacts of global warming
- 2.2.6 Sector-based impacts assessment of climate system tipping points
- 2.3 Negative social tipping points
- 2.4 Cascades of tipping in impacts
- 2.5 Early warning of tipping points in impacts
- References
- Section 3
-
-
Considers how to govern Earth system tipping points and their associated risks. We look at governance of mitigation, prevention and stabilisation then we focus on governance of impacts, including adaptation, vulnerability and loss and damage. Finally, we assess the need for knowledge generation at the science-policy interface.
-
-
- 3.1 Introduction
- 3.2 Prevention of Earth system tipping processes
- 3.3 Tipping point impact governance
- 3.4 Knowledge co-production and science-policy engagement
- References
-
-
-
- 3. Governance of Earth system tipping points
- 3.1 Governing Earth system tipping points – Introduction
- 3.2 Prevention of Earth system tipping processes
- 3.3 Tipping point impact governance
- 3.4 Knowledge co-production and science-policy engagement
- References
- Section 4
-
-
Focuses on positive tipping points in technology, the economy and society. It provides a framework for understanding and acting on positive tipping points. We highlight illustrative case studies across energy, food and transport and mobility systems, with a focus on demand-side solutions (which have previously received limited attention).
-
-
- 4.1 Introduction
- 4.2 Understanding and acting on positive tipping points
- 4.3 Positive tipping points in energy, transport and food systems
- 4.3.1 Energy systems
- 4.3.2 Transport and mobility systems
- 4.3.3 Food systems
- 4.3.3.1 Introduction
- 4.3.3.2 Avoiding food loss and waste
- 4.3.3.3 Shifting towards more plant-based diets
- 4.3.3.4 Improving alternatives to animal products
- 4.3.3.5 Shifting Farming Practice
- 4.3.3.6 Food system tipping points have important feedbacks for protecting nature
- 4.3.3.7 Strategic interventions to enable positive tipping points in food systems
- 4.4 Cross-cutting enablers of positive tipping points
- 4.4.1 Socio-behavioural systems
- 4.4.2 Political systems
- 4.4.2.1 Introduction
- 4.4.2.2 Political systems can enable (or dampen) social tipping
- 4.4.2.3 Political systems themselves can tip
- 4.4.2.4 Civil society & political tipping: The role of social movements and coalition formation
- 4.4.2.5 International climate governance and the diffusion of political change
- 4.4.3. Financial systems
- 4.4.4 Digitalisation
- 4.4.4.1 Introduction
- 4.4.4.2 Conceptual underpinnings
- 4.4.4.3 Digital technologies and avoid options: Teleworking
- 4.4.4.4 Digital technologies and shift options: Mobility-as-a-Service
- 4.4.4.5 Digital technologies and improve options: Smart homes
- 4.4.4.6 Other domains where digital technologies can enable positive tipping
- 4.4.4.7 Strategic interventions
- 4.4.5 Detecting ‘early opportunity indicators’ for positive tipping points
- 4.4.5.1 Predicting tipping points
- 4.4.5.2 Detecting early opportunity signals indicators in the electric vehicle transition
- 4.4.5.3 Resilience change prior to the EV transition tipping point
- 4.4.5.4 Changes detectable in other social data?
- 4.4.5.5 Limitations
- 4.4.5.6 Measuring progress – Early opportunity indicators in other sectors
- 4.5 Positive tipping cascades
- 4.6 Risks, equity and justice in the governance of positive tipping points
- References
-
-
-
- 4. Positive tipping points in technology, economy and society
- 4.1 Introduction
- 4.2 Understanding and acting on positive tipping points
- 4.3 Positive tipping points in energy, transport and food systems
- 4.3.1 Energy systems
- 4.3.2 Transport and mobility systems
- 4.3.3 Food systems
- 4.3.3.1 Introduction
- 4.3.3.2 Avoiding food loss and waste
- 4.3.3.3 Shifting towards more plant-based diets
- 4.3.3.4 Improving alternatives to animal products
- 4.3.3.5 Shifting Farming Practice
- 4.3.3.6 Food system tipping points have important feedbacks for protecting nature
- 4.3.3.7 Strategic interventions to enable positive tipping points in food systems
- 4.4 Cross-cutting enablers of positive tipping points
- 4.4.1 Socio-behavioural systems
- 4.4.2 Political systems
- 4.4.2.1 Introduction
- 4.4.2.2 Political systems can enable (or dampen) social tipping
- 4.4.2.3 Political systems themselves can tip
- 4.4.2.4 Civil society & political tipping: The role of social movements and coalition formation
- 4.4.2.5 International climate governance and the diffusion of political change
- 4.4.3. Financial systems
- 4.4.4 Digitalisation
- 4.4.4.1 Introduction
- 4.4.4.2 Conceptual underpinnings
- 4.4.4.3 Digital technologies and avoid options: Teleworking
- 4.4.4.4 Digital technologies and shift options: Mobility-as-a-Service
- 4.4.4.5 Digital technologies and improve options: Smart homes
- 4.4.4.6 Other domains where digital technologies can enable positive tipping
- 4.4.4.7 Strategic interventions
- 4.4.5. Detecting ‘early opportunity indicators’ for positive tipping points
- 4.4.5.1 Predicting tipping points
- 4.4.5.2 Detecting early opportunity signals indicators in the electric vehicle transition
- 4.4.5.3 Resilience change prior to the EV transition tipping point
- 4.4.5.4 Changes detectable in other social data?
- 4.4.5.5 Limitations
- 4.4.5.6 Measuring progress – Early opportunity indicators in other sectors
- 4.5 Positive tipping cascades
- 4.6 Risks, equity and justice in the governance of positive tipping points
- References
- Appendices






